

 Реферат: Кариотип человека
Реферат: Кариотип человека
В полном соответствии с данными общей цитогенетики о слабом отрицательном влиянии дисбаланса по гетерохроматиновому материалу на развитие организма находятся сведения о существовании в человеческой популяции значительного полиморфизма, обусловленного размерами околоцентромерного гетерохроматина. Особенно сильно варьирует содержание структурного гетерохроматина С-типа в аутосомах 1, 4, 9, 13—15, 16, 21—22 и Y-хромосоме. Отсутствие фенотипических отклонений от нормы у большинства носителей таких кариотипических вариантов позволяет рассматривать их как варианты нормы. Однако эта проблема поставлена на повестку дня совсем недавно. Она требует тщательных исследований на большом популяционном материале, прежде чем будут намечены обоснованные границы хромосомной нормы, за пределами которой для организма становится не безразличным дисбаланс и по гетерохроматину.
Есть много оснований рассматривать хромосомные районы, положительно окрашивающиеся по G-методике, как разновидность структурного гетерохроматина. В пользу этого представления, помимо отношения к красителям, свидетельствуют поздняя репликация этих районов, образование ими хромомер в профазных мейотических хромосомах, способность отставать в митотической конденсации под влиянием 5-бромдезоксиуридина или холода. Важно отметить, что дисбаланс по аутосомам, особенно богатым G-окрашивающимся хроматином, влечет за собой возникновение наименее тяжелых аномалий развития для индивида — носителя такого дисбаланса. Так, именно к этой категории хромосомных аномалий относятся трисомии 13, 18 и 21. Имеются сообщения и о том, что ДНК со средней повторяемостью одинаковых нуклеотидных последовательностей локализуется в G-окрашивающихся сегментах хромосом.
Вопросы, которые стоят перед цитогенетикой человека в отношении структуры, локализации и особенно генетического значения структурного гетерохроматина, сравнительно новые.
Прогресс в их разрешении нельзя отделить от прогресса в расшифровке природы гетерохроматина у эукариотов в целом.
Помимо структурного гетерохроматина, существует ф а-культативный гетерохроматин, появление которого в хромосоме обусловлено гетерохроматинизацией эухроматических районов при особых условиях. Имеются достоверные доказательства существования этого явления в хромосомах человека на примере генетической инактивации одной из Х-хромосом в соматических клетках женщины. У человека и других млекопитающих это частный случай явления, впервые открытого на дрозофиле Muller в 1932 г. и получившего название «компенсации дозы гена». Для млекопитающих его сущность состоит в эволюционно сформировавшемся механизме инактивации второй дозы генов, локализованных в Х-хромосоме, благодаря чему, несмотря на неодинаковое число Х-хромосом, мужской и женский организмы по количеству функционирующих генов уравнены.
Сформулированная Lyon (1961, 1974) соответствующая гипотеза, получившая ее имя, состоит из трех основных положений:
1. В соматических клетках нормального женского организма одна из двух Х-хромосом инактивирована.
2. В разных клетках организма инактивируется или материнская, или отцовская Х-хромосома.
3. Инактивация происходит в раннем эмбриональном периоде и стойко сохраняется за данной Х-хромосомой в клеточных поколениях.
Гипотеза Lyon основана на большом числе генетических и цитологических фактов, в том числе полученных на человеке, которые за годы с момента ее выдвижения непрерывно пополнялись и сведения о которых можно найти в ряде обзоров (А. Ф. Захаров, 1968; Lyon, 1972, 1974; Ghan-dra, Brown, 1975, и др.).
Генетические факты основаны на том, что у гетерозигот по сцепленным с Х-хромосомой признакам обнаруживаются две клеточные популяции. В одной из них проявляется действие гена материнской Х-хромосомы, в другой — отцовской, что связано с инактивацией отцовского или материнского аллелей соответственно. При формулировании своей гипотезы Lyon опиралась на случаи мозаичной окраски шерстного покрова мышей, что обусловливалось инактивацией в разных участках тела либо дикого гена, либо его мутантного аллеля. У человека обстоятельные доказательства существования в организме гетерозиготных женщин двух популяций клеток, в каждой из которых инактивирован один из двух аллелей гена, локализованного в Х-хромосоме, получены при изучении эффектов генов глюкозо-6-фосфатдегидрогеназы, фосфоглицераткиназы, гипоксантин-фосфорибозилтрансферазы, эритроци-тарной группы крови Xg (а), при изучении сцепленных с Х-хромосомой агаммаглобулинемии и мукополисахаридоза (синдром Хантера), гемофилии. У гетерозигот по электро-форетическим вариантам глюкозо-6-фосфатдегидрогеназы подтверждено, что у человека Х-хромосома инактивируется в раннем эмбриональном периоде (Migeon, Kennedy, 1975). Эти выводы необходимо иметь в виду при интерпретации данных по наследственным болезням, сцепленным с Х-хромосомой, особенно у монозиготных близнецов.
Цитологические доказательства в пользу гипотезы Lyon также весьма убедительны и состоят в том, что в нормальных женских соматических клетках одна из двух Х-хромосом отвечает характеристикам гетерохроматинизированной хромосомы. В интерфазном ядре она обнаруживается в виде так называемого тельца Барра (Х-хроматина) — плотно конденсированной, интенсивно окрашивающейся глыбки хроматина. В профазе эта хромосома опережает в цикле конденсации своего гомолога — вторую Х-хромосому. В условиях экспериментального воздействия холодом или 5-бромдезоксиуридином одна из Х-хромосом значительно отстает в конденсации, не отличаясь в этом отношении от структурного гетерохроматина аутосом 1, 9, 16 и Y-хромосомы. Вторая Х-хромосома является одной из наиболее запаздывающих по началу и окончанию репликации ДНК.
Исследование многочисленных случаев аномалий в системе Х-хромосом у человека показывает, что явление компенсации дозы генов распространяется также на все случаи нарушений в числе Х-хромосом, оставляя в соматической клетке лишь одну Х-хромосому в активном состоянии. Особенно демонстративны в этом отношении Х-полисомии, когда число инактивированных Х-хромосом равно числу имеющихся в клетке за вычетом одной генетически функционирующей.
Как было показано выше, сведения о кариотипе человека постоянно углубляются, и исследования все больше Проводятся на молекулярном уровне. Цитологическое изучение материальных основ наследственности человека хорошо дополняется генетическим анализом дискретных признаков.
Глава 3. Цитогенетический метод.
В генетике человека используются разнообразные методы исследования, применяемые и в других разделах биологии — генетике, физиологии, цитологии, биохимии и др. Антропогенетика располагает также собственными методами исследования: цитогенетическим, близнецовым, генеалогическим и др.[4]
Достижениями молекулярной биологии и биохимии внесен большой вклад в развитие генетики. В настоящее время биохимическим и молекулярно-генетическим методам исследования принадлежит ведущая роль в генетике человека и медицинской генетике. Однако и классические методы генетики человека, такие как цитогенетический, генеалогический и близнецовый, имеют существенное значение в настоящее время, особенно в вопросах диагностики, медико-генетического консультирования и прогнозирования потомства.
Ознакомимся с возможностями цитогенетического метода.
Суть этого метода заключается в изучении строения отдельных хромосом, а также особенностей набора хромосом клеток человека в норме и патологии. Удобным объектом для этого служат лимфоциты, клетки эпителия щеки и другие клетки, которые легко получать, культивировать и подвергать кариологическому анализу. Это важный метод определения пола и хромосомных наследственных заболеваний человека.
Основой цитогенетического метода является изучение морфологии отдельных хромосом клеток человека. Современный этап познания строения хромосом характеризуется созданием молекулярных моделей этих важнейших структур ядра, изучением роли отдельных компонентов хромосом в хранении и передаче наследственной информации.
В главе 1 мы рассмотрели такие компоненты хромосом, как белки и нуклеиновые кислоты. Здесь же кратко остановимся на строении и морфологии хромосом.
Строение хромосом.
Хромосомную теорию наследственности создал американский ученый Т. Г. Морган. Проведя большое количество исследований на плодовой мушке дрозофиле, Морган и его ученики установили, что именно в хромосомах находятся открытые Менделем факторы наследственности, которые были названы генами. Т. Морган и его ученики показали, что гены расположены линейно по длине хромосомы.
После того как было доказано, что хромосомы являются основными генофорами (носителями генов), начался период их наиболее интенсивного изучения. Успехи молекулярной биологии и генетики позволили понять некоторые закономерности строения и функционирования хромосом прокариот и эукариот, однако многое здесь остается еще неизвестным. В последние годы хромосомы эукариот, особенно человека, становятся предметом изучения различных специалистов, начиная от генетиков и кончая физиками.
 |
В настоящее время установлено, что в основе строения хромосомы лежит хроматин — сложный комплекс ДНК, белков, РНК и других веществ, входящих в хромосому (строение хроматина мы подробно рассмотрели в главе 1). Предполагается, что в хромосому человека входит одна гигантская молекула ДНК, молекулы РНК, гистоны и кислые белки, различные ферменты, фосфолипиды, металлы Са2+, Mg2+ и некоторые другие вещества. Способ укладки и взаимного расположения молекул этих химических соединений в хромосоме пока не известен. Длинная нить ДНК не может располагаться в хромосоме беспорядочно. Существует предположение, что нить ДНК упакована закономерным образом и связана с белками.
![]()
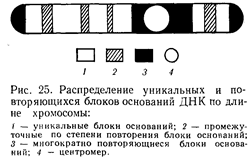
Ф. Арриги и соавторы (1971) установили, что уникальные последовательности занимают более 56% ДНК хромосом человека, высокоповторяющиеся — 12,4 %, промежуточные повторы — 8 %. Общее количество повторяющихся генов в ДНК хромосомы человека равно 28%. Число хромосом у человека длительное время оставалось невыясненным. Дело в том, что определить количество хромосом у млекопитающих, особенно у человека, было трудно. Хромосомы оказались маленькими, весьма многочисленными, плохо поддавались подсчету. При фиксации клетки они сливались в комки, что затрудняло определение истинного числа хромосом. Поэтому первые исследователи не могли точно и правильно подсчитать количество хромосом в клетках человека. Называлось разное количество хромосом — от 44 до 50.
![]()
 Обычно хромосомы в клетках наблюдают
во время митоза на стадии метафазной пластинки. В интерфазном ядре хромосомы в
световой микроскоп не видны. В 1912 г. Г. Винивартер, изучая хромосомы в
сперматогониях и оогониях половых желез человека, удаленных во время операции,
установил, что мужской набор хромосом (кариотип) содержит 47 хромосом, а
женский — 48. В 1922 г. Т. Пайнтер повторил исследования Винивартера и
установил, что мужской и женский кариотипы содержат по 48 хромосом, но женский
отличается от мужского только двумя хромосомами. У женщин находится 2 большие
половые хромосомы, а у мужчины одна большая Х-хромосома и одна маленькая
К-хромосома. В последующие годы эту точку зрения поддерживали и другие ученые.
П. И. Живаго и А. Г. Андреа (1932) предложили первую классификацию хромосом в
зависимости от их длины. Так как хромосомы очень близко располагаются одна
около другой и их очень трудно исследовать, то и в последующие годы точное
число хромосом у человека служило предметом споров и дискуссий. Однако
постепенно было достигнуто согласие между исследователями по этому вопросу, и в
течение 30 лет большинство цитогенетиков считало, что у человека диплоидное
число хромосом равно 48, а гаплоидное — 24. Усовершенствованные методы изучения
хромосом позволили получить более точные сведения о количестве хромосом в
клетках у человека, а также выявить аномалии нормального кариотипа,
ответственные за некоторые уродства. Особенно плодотворным оказались два
метода:
Обычно хромосомы в клетках наблюдают
во время митоза на стадии метафазной пластинки. В интерфазном ядре хромосомы в
световой микроскоп не видны. В 1912 г. Г. Винивартер, изучая хромосомы в
сперматогониях и оогониях половых желез человека, удаленных во время операции,
установил, что мужской набор хромосом (кариотип) содержит 47 хромосом, а
женский — 48. В 1922 г. Т. Пайнтер повторил исследования Винивартера и
установил, что мужской и женский кариотипы содержат по 48 хромосом, но женский
отличается от мужского только двумя хромосомами. У женщин находится 2 большие
половые хромосомы, а у мужчины одна большая Х-хромосома и одна маленькая
К-хромосома. В последующие годы эту точку зрения поддерживали и другие ученые.
П. И. Живаго и А. Г. Андреа (1932) предложили первую классификацию хромосом в
зависимости от их длины. Так как хромосомы очень близко располагаются одна
около другой и их очень трудно исследовать, то и в последующие годы точное
число хромосом у человека служило предметом споров и дискуссий. Однако
постепенно было достигнуто согласие между исследователями по этому вопросу, и в
течение 30 лет большинство цитогенетиков считало, что у человека диплоидное
число хромосом равно 48, а гаплоидное — 24. Усовершенствованные методы изучения
хромосом позволили получить более точные сведения о количестве хромосом в
клетках у человека, а также выявить аномалии нормального кариотипа,
ответственные за некоторые уродства. Особенно плодотворным оказались два
метода:
1. Обработка культуры клеток алкалоидом колхицином, который ведет к накоплению делящихся клеток на стадии метафазы;
2. Обработка клеток слабыми растворами солей, вызывающими набухание, расправление хромосом, что облегчает их исследование.
В 1956 г. шведские цитологи Дж. Тийо и А. Леван изготовили культуры клеток из тканей легких, взятых у абортированных человеческих эмбрионов и, используя усовершенствованную методику обработки клеток, получили необычайно четкие препараты, в которых ясно было видно 46 хромосом.[5]
Несколькими месяцами позднее Ч. Форд и Дж. Хаммертон в Англии установили, что диплоидные предшественники половых клеток в семенниках мужчин (сперматогонии) также имеют по 46 хромосом, а гаплоидные (сперматоциты 1-го деления) — по 23 хромосомы.
После этого были изучены многие клетки из разных органов и тканей человека и везде нормальное число хромосом оказалось равным 46.
Женский кариотип отличается от мужского только одной половой хромосомой. Остальные 22 пары одинаковы у мужчин и женщин. Эти 22 пары хромосом называются аутосомами. Нормальный кариотип состоит из 44 аутосом (22 пары) и двух половых хромосом — XX у женщин и XY у мужчин, т. е. женский кариотип имеет две большие половые хромосомы, а мужской — одну большую и одну маленькую.
В половых клетках человека находится одинарный (гаплоидный) набор хромосом — 23, а в соматических клетках — двойной (диплоидный) набор — 46. Эти открытия стимулировали дальнейшее изучение хромосом. Были разработаны методы исследования хромосом в культуре лимфоцитов периферической крови и на других объектах. В настоящее время хромосомы относительно легко исследуют в лимфоцитах периферической крови. Венозную кровь помещают в специальную питательную среду, добавляют фитогемаглютинин, который стимулирует клетки к делению, и помещают на 72 ч. в термостат. За 6 ч. до конца инкубации сюда добавляют колхицин, который задерживает процесс деления клеток на стадии метафазной пластинки. Затем культуру помещают в гипотонический раствор NaCl, в котором клетки набухают, что приводит к легкому разрыву оболочек ядра и переходу хромосом в цитоплазму. После этого препараты окрашивают ядерными красителями, в частности ацетоорсеином, и рассматривают их в световом микроскопе с иммерсией.
Под микроскопом учитывают общее количество хромосом, фотографируют их, затем из фото вырезают ножницами каждую хромосому и наклеивают на чистый лист бумаги в ряд, начиная от самой большой (первой) хромосомы и кончая самой маленькой (двадцать второй) и половой Y-хромосомой. Люминесцентная методика позволяет быстро и просто проводить массовые исследования с целью выявления больных с различными типами хромосомных аномалий. Совокупность количественных (число хромосом и их размеры) и качественных (морфология хромосом) признаков диплоидного набора единичной клетки обозначается термином «кариотип». Строение хромосом изменяется в зависимости от стадии деления клеток (профазы, метафазы, анафазы, телофазы).
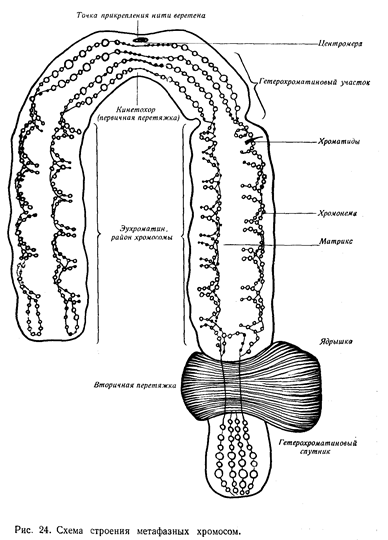
Уже в профазе митоза видно, что хромосома образована двумя взаимно переплетающимися нитями одинакового диаметра — хроматидами. В метафазе хромосома уже спирализована, и две ее хроматиды ложатся параллельно, разделенные узкой щелью. Каждая хроматида состоит из двух полухроматид. В результате митоза хроматиды материнской хромосомы становятся сестринскими хромосомами, а полухроматиды — их хроматидами. В основе хроматид лежат хромонемы — так называют более тонкие нити ДНП, состоящие из белка и нуклеиновых кислот.
В интерфазе (промежуток между двумя
делениями клеток) хроматин тесно связан с ядерными мембранами и ядерным
белковым матриксом. Он образует также большие участки деспирализованных нитей
ДНП. Затем постепенно хроматин спирализуется, образуя типичные метафазные 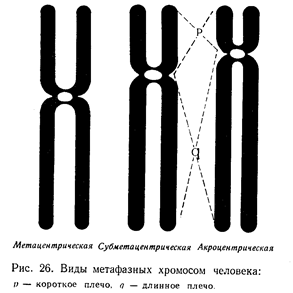 хромосомы. Размеры их
варьируют от 2 до 10 микрон.
хромосомы. Размеры их
варьируют от 2 до 10 микрон.
В настоящее время интенсивно исследуются структурные особенности аутосом и половых хромосом (на клетках костного мозга, лимфоцитах, фибробластах, клетках кожи, регенерирующей печени).
