

 Реферат: Кардиография (введение )
Реферат: Кардиография (введение )
Мониторное электрокардиографическое наблюдение наиболее часто используют при остром инфаркте миокарда. Его проводят обычно в отделениях или палатах интенсивной терапии в первые дни после возникновения инфаркта, при наличии преходящих нарушений ритма и проводимости, которые требуют срочных терапевтических мероприятий, а также для уточнения диагноза аритмии. Кроме того, его используют иногда при проведении массивной противоаритмической или сердечной терапии, а также при применении отдельных диагностических процедур, которые могут приводить к возникновению аритмий (например, проба с физической нагрузкой, зондирование сердца, ангиокардиография и т. д.). Нередко ЭКГ записывают на магнитную ленту, что позволяет вводить и анализировать ЭКГ с помощью ЭВМ.
Современная медицина базируется на широком использовании разнообразной аппаратуры, которая в большинстве своем является физической по конструкции. Поэтому в курсе медицинской и биологической физике рассматриваются устройство и принципы работы основной медицинской аппаратуры.
2. БИОЭЛЕКТРИЧЕСКИЕ ОСНОВЫ ЭЛЕКТРОКАРДИОГРАФИИ
2.1 МЕМБРАННАЯ ТЕОРИЯ ВОЗНИКНОВЕНИЯ БИОПОТЕНЦИАЛОВ
В основе возникновения электрических явлений в сердце лежит, как известно, проникновение ионов калия (К+), натрия (Na+), кальция (Са 2+), хлора (СГ) и др. через мембрану мышечной клетки. В электрохимическом отношении клеточная мембрана представляет собой оболочку, обладающую разной проницаемостью для различных ионов. Она как бы разделяет два раствора электролитов, существенно отличающихся по своему составу. Внутри клетки, находящейся в невозбужденном состоянии, концентрация К+ в 30 раз выше, чем во внеклеточной жидкости. Наоборот, во внеклеточной среде примерно в 20 раз выше концентрация Na+, в 13 раз выше концентрация СГ и в 25 раз выше концентрация Са2+ по сравнению с внутриклеточной средой. Такие высокие градиенты концентрации ионов по обе стороны мембраны поддерживаются благодаря функционированию в ней ионных насосов, с помощью которых ионы Na, Ca и Сl выводятся из клетки, а ионы К входят внутрь клетки. Этот процесс осуществляется против концентрационных градиентов этих ионов и требует затраты энергии.
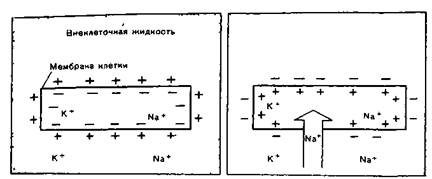
А Б
Клетка миокарда в покое (А) и во время деполяризации (Б).
В невозбужденной клетке мембрана более проницаема для К+ и СГ. Поэтому ионы К+ в силу концентрационного градиента стремятся выйти из клетки, перенося свой положительный заряд во внеклеточную среду. Ионы СГ, наоборот, входят внутрь клетки, увеличивая тем самым отрицательный заряд внутриклеточной жидкости. Это перемещение ионов и приводит к поляризации клеточной мембраны невозбужденной клетки: наружная ее поверхность становится положительной, а внутренняя - отрицательной. Возникающая таким образом на мембране разность потенциалов препятствует дальнейшему перемещению ионов (К - из клетки и С1 - в клетку), и наступает стабильное состояние поляризации мембраны клеток сократительного миокарда в период диастолы. Если мы теперь с помощью микроэлектродов измерим разность потенциалов между наружной и внутренней поверхностью клеточной мембраны, то зарегистрируем так называемый трансмембранный потенциал покоя (ТМПП), имеющий отрицательную величину, в норме составляющую около - 90 mV.
При возбуждении клетки резко изменяется проницаемость ее стенки по отношению к ионам различных типов. Это приводит к изменению ионных потоков через клеточную мембрану и, следовательно, к изменению величины самого ТМПП. Кривая изменения трансмембранного потенциала во время возбуждения получила название трансмембранного потенциала действия (ТМПД). Различают несколько фаз ТМПД миокардинальной клетки (рисунок 1).
Фаза 0. Во время этой начальной фазы возбуждения - фазы деполяризации - резко увеличивается проницаемость мембраны клетки для ионов Na, которые быстро устремляются внутрь клетки (быстрый натриевый ток). При этом, естественно, меняется заряд мембраны: внутренняя поверхность мембраны становится положительной, а наружная - отрицательной. Величина ТМПД изменяется от -90 mV до +20 mV, т.е. происходит реверсия заряда - перезарядка мембраны. Продолжительность этой фазы не превышает 10 мс.
Фаза 1. (фаза начальной быстрой реполяризации) Как только величина ТМПД достигает примерно +20 mV, проницаемость мембраны для Na+ уменьшается, а для СГ. Это приводит к возникновению небольшого тока отрицательных ионов С1 внутри клетки, которые частично нейтрализуют избыток положительных ионов Na внутри клетки, что ведет к некоторому падению ТМПД примерно до 0 или ниже.
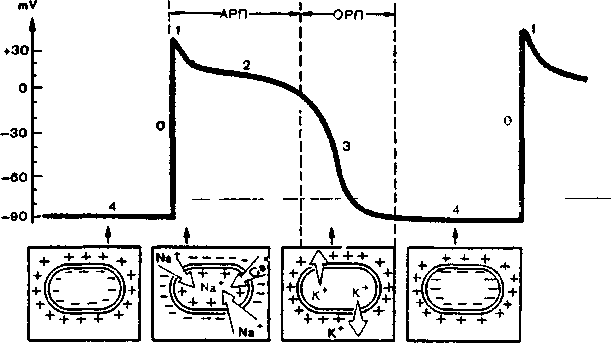
Рисунок 1. Трансмембранный потенциал действия (ТМПД). АРП и ОРП - абсолютный и относительный рефракторный периоды.
Фаза 2. (фаза плато) В течение этой фазы величина ТМПД поддерживается примерно на одном уровне, что приводит к формированию на кривой ТМПД своеобразного плато. Постоянный уровень величины ТМПД поддерживается при этом за счет медленного входящего тока Са2+ и Na+ направленного внутрь клетки, и тока К+ из клетки. Продолжительность этой фазы велика и составляет около 200 мс. В течение фазы 2 мышечная клетка остается в возбужденном состоянии, начало ее характеризуется деполяризацией, окончание - реполяризацией мембраны.
Фаза 3. (конечной быстрой реполяризации) К началу фазы 3 резко уменьшается проницаемость клеточной мембраны для Na+ и Са2+ и значительно возрастает проницаемость ее для К+. Поэтому вновь начинает преобладать перемещение ионов К наружу из клетки, что приводит к восстановлению прежней поляризации клеточной мембраны, имевшей место в состоянии покоя: наружная ее поверхность вновь оказывается заряженной положительно, а внутренняя поверхность - отрицательно. ТМПД достигает величины ТМПП.
Фаза 4. (фаза диастолы) Во время этой фазы ТМПД происходит восстановление исходной концентрации К+, Na+, Ca2+, СГ соответственно внутри и вне клетки благодаря действию «Na+ - K+ -насоса». При этом уровень ТМПД мышечных клеток остается на уровне примерно - 90 mV.
Клетки проводящей системы сердца и клетки синусового узла обладают способностью к спонтанному медленному увеличению ТМПП - уменьшению отрицательного заряда внутренней поверхности мембраны во время фазы 4. Этот процесс получил название спонтанной диастолической деполяризации и лежит в основе автоматической активности клеток синоатриального (синусового) узла и проводящей системы сердца, т. е. способности к «самопроизвольному» зарождению в них электрического импульса.
Наружная поверхность клеточной мембраны заряжена:
1) положительно
- в невозбужденной мышечной клетке, находящейся
в состоянии покоя;
2) отрицательно - в клетке, находящейся в состоянии возбуждения
в
фазе 0 и 1 ТМПД (деполяризация и ранняя быстрая
реполяризация);
3) положительно
- в клетке, восстанавливающей свой исходный
потенциал (реполяризация клетки).
2.2 ОСНОВНЫЕ ФУНКЦИИ СЕРДЦА
Сердце обладает рядом функций, определяющих особенности его работы.
1) Функция автоматизма
Функция автоматизма заключается в способности сердца вырабатывать электрические импульсы при отсутствии внешних раздражений.
Функцией автоматизма обладают клетки синоатриального узла (СА-узла) и проводящей системы сердца: атриовентрикулярного соединения (АВ-соединения), проводящей системы предсердий и желудочков. Они получили название клеток водителей /пейсмекеров (от англ., pacemaker— водитель). Сократительный миокард лишен функции автоматизма.
Если в норме ТМПД
сократительных мышечных клеток в течение всей диастолической
фазы (фазы 4 ТМПД) стабильно поддерживается на одном и том же уровне, равном примерно-90 mV, то для волокон водителей
ритма (пейсмекеров) характерно медленное спонтанное
уменьшение
мембранного потенциала в диастолу, как это показано на
рисунке 2. Этот
процесс носит название медленной спонтанной диастолической деполяризации и
возникает в результате особых свойств мембраны пейсмекеров - постепенного
самопроизвольного увеличения
в диастолу проницаемости мембраны для ионов Na, медленно входящих в клетку. В результате скопления в клетке
все большего количества положительных ионов
отрицательный заряд внутренней поверхности
клеточной мембраны частично нейтрализуется, и разность потенциалов между наружной и внутренней поверхностью
мембраны (ТМПП) постепенно уменьшается. Как только ТМПП достигнет критического уровня (примерно 60 mV)9 проницаемость
мембраны для ионов Na резко и быстро возрастает,
что приводит к возникновению быстрой лавинообразной деполяризации клетки (фаза
О ТМПД) - ее возбуждению, которая является
импульсом к возбуждению других клеток миокарда. Критический потенциал покоя
|
|
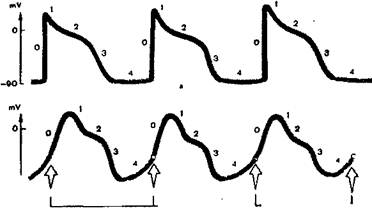
Рисунок 2. Спонтанная диастолическая деполяризация волокон водителей ритма - пейсмекеров. а) - ТМПД мышечных клеток; б) - ТМПЛ клеток пейсмекеров.
Понятно, что чем выше скорость спонтанной диастолической деполяризации, тем чаще в клетках водителя ритма возникают электрические импульсы. В норме максимальной скоростью диастолической деполяризации и максимальной автоматической активностью обладают клетки СА-узла, который вырабатывает электрические импульсы с частотой около 60 -80 в минуту. Это центр автоматизма первого порядка.
Функцией автоматизма обладают некоторые участки в предсердиях и АВ-соединение зона перехода атриовентрикулярного узла (АВ-узла) в пучок Гиса (по международной анатомической номенклатуре - предсердно-желудочковый пучок)
Эти участки проводящей системы сердца, являющиеся центрами автоматизма второго порядка, могут продуцировать электрические импульсы с частотой 40-60 в минуту. Следует подчеркнуть, что сам АВ-узел, также входящий в состав АВ-соединения, не обладает функцией автоматизма.
Межпредсердный пучок (Бахмана)
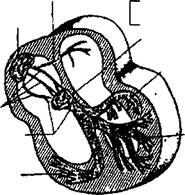
| Левая передняя ветвь пучка Гиса |
Правая ножка пучка Гиса
| левая задняя ветвь пучка Гиса |
| АВ-узел |
| Межузловые проводящие тракты (Бахмана Венкебаха, Тореля) |
АВ-соединение
Правая ножка пучка Гиса
Рисунок 3. Проводящая система сердца
Наконец, центрами автоматизма третьего порядка, обладающими самой низкой способностью к автоматизму (25-45 импульсов в минуту), являются нижняя часть пучка Гиса, его ветви и волокна Пуркинье. Однако в норме возбуждение сердца происходит только в результате импульсов, возникающих в волокнах СА-узла, который является единственным нормальным водителем ритма. Дело в том, что в условиях сравнительно частой им-пульсации СА-узла подавляется автоматизм клеток АВ-соединения, пучка Гиса и волокон Пуркинье. Последние являются только потенциальными, или латентными, водителями ритма. При поражениях СА-узла функцию водителя ритма могут взять на себя нижележащие отделы проводящей системы сердца - центры автоматизма II и даже III порядка.
1. Все
волокна проводящей сметены сердца (кроме средней части
АВ-узла) потенциально обладают функцией автоматизма.
2. В
норме единственным водителем ритма является СА-узел,
который подавляет автоматическую активность остальных
(эктопических) водителей ритма сердца.
На функцию СА-узла и других водителей ритма большое влияние оказывает симпатическая и парасимпатическая нервная система: активизация симпатической системы ведет к увеличению автоматизма клеток СА-узла и проводящей системы, а парасимпатической системы - к уменьшению их автоматизма.
2) Функция проводимости
Функция проводимости - это способность к проведению возбужде-ния, возникшего в каком-либо участке сердца, к другим отделам сердечной мышцы.
Функцией проводимости обладают как волокна специализированной проводящей системы сердца, так и сократительный миокард; однако в последнем случае скорость проведения электрического импульса значительно меньше.
Следует хорошо усвоить последовательность и особенности распространения возбуждения по различным отделам проводящей системы сердца. В норме волна возбуждения, генерированного в клетках СА-узла, распространяется по короткому проводящему пути на правое предсердие, по трем межузловым трактам - Бахмана, Венкебаха и Тореля - к АВ-узлу и по межпредсердному пучку Бахмана - на левое предсердие. Возбуждение распространяется по этим проводящим трактам в 2-3 раза быстрее, чем по миокарду предсердий. Общее направление движения волны возбуждения - сверху вниз и несколько влево от области СА-узла к верхней части АВ-узла. Вначале возбуждается правое предсердие, затем присоединяется левое, в конце возбуждается только левое предсердие (рисунок 4). Скорость распространения возбуждения здесь невелика и составляет в среднем около 30 - 80 см-с"1. Время охвата волной возбуждения обоих предсердий не превышает 0,1 с.
1. Направление распространения волны возбуждения по предсердиям - сверху вниз и немного влево.
2. Вначале возбуждается правое, затем правое и левое предсердия, в конце - только левое предсердие.
3 Время охвата возбуждением предсердий не превышает в норме 0,1 с.
В АВ-узле и особенно в пограничных участках между АВ-узлом и пучком Гиса происходит значительная задержка волны возбуждения, скорость проведения не более2-5 см с". Задержка возбуждения в АВ-узле способствует тому, что желудочки начинают возбуждаться только после окончания полноценного сокращения предсердий и желудочков. Малая скорость проведения электрического импульса в АВ-узле обусловливает и другую особенность его функционирования: АВ-узел может «пропустить» из предсердий в желудочки не более 180 - 200 импульсов в минуту. Поэтому при учащении сердечного ритма более 180 - 200 ударов в минуту некоторые импульсы из предсердий не достигают желудочков, наступает так называемая атриовентрикулярная блокада проведения. В этом отношении АВ-узел является одним из самых уязвимых отделов проводящей системы сердца
1. В АВ - узле происходит физиологическая задержка волны возбуждения, определяющая нормальную временную последовательность возбуждения предсердий и желудочков.
2. При учащении сердечных импульсов, исходящих из СА-узла или предсердий, бол eel 80-220 в минуту, даже у здорового человека может наступить частичная (атриовентрикулярная) блокада проведения электрического импульса от предсердий к желудочкам. От АВ-узла волна возбуждения передается на хорошо развитую внутрижелудочковую проводящую систему, состоящую из предсердно-желудочкового пучка (пучка Гиса), основных ветвей (ножек) пучка Гиса и волокон Пуркинье.
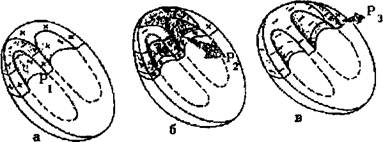
Рисунок 4. Распространение возбуждения по предсердиям. а) - начальное возбуждение правого предсердия; б) - возбуждение правого и левого предсердий; в) конечное возбуждение левого предсердия. Красным цветом показаны возбужденные (заштрихованные) и возбуждающиеся в настоящий момент (сплошные) участки Р1 ,Р2, РЗ - моментные векторы деполяризации предсердий.
В норме скорость проведения по пучку Гиса и его ветвям составляет 100 -150 см-с ", а по волокнам Пуркинье -300 - 400 см-с "!. Большая скорость проведения электрического импульса по проводящей системе желудочков способствует почти одновременному охвату желудочков волной возбуждения и наиболее оптимальному и эффективному выбросу крови в аорту и легочную артерию. В норме общая продолжительность деполяризации желудочков колеблется от 0,06 до 0,10 с.
