

 Реферат: Роль материнского генома в развитии потомка
Реферат: Роль материнского генома в развитии потомка
Помимо нуклеиновых кислот и белков для построения новых митохондрий нужны липиды. В отличие от хлоропластов митохондрии получают бóльшую часть своих липидов извне. В животных клетках фосфолипиды, синтезированные в эндоплазматическом ретикулуме, транспортируются к наружной мембране митохондрий с помощью особых белков, а затем включаются во внутреннюю мембрану; как полагают, это происходит в месте контакта двух мембран. Основная реакция биосинтеза липидов, катализируемая самими митохондриями, - это превращение фосфатидной кислоты в фосфолипид кардиолипин, который содержится главным образом во внутренней митохондриальной мембране и составляет около 20% всех ее липидов.
Размеры и форма митохондриальных геномов
К настоящему времени прочитано более 100 разных геномов митохондрий. На-бор и количество их генов в митохондриальных ДНК, для которых полностью определена последовательность нуклеотидов, сильно различаются у разных ви-дов животных, растений, грибов и простейших. Наибольшее количество генов обнаружено в митохондриальном геноме жгутикового простейшего Rectinomo-nas americana — 97 генов, включая все кодирующие белок гены, найденные в мтДНК других организмов. У большинства высших животных геном митохон-дрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и два для рРНК (для большой субъединицы рибосом 16S рРНК и для малой 12S рРНК). У растений и простейших, в отличие от животных и большинства гри-бов, в митохондриальном геноме закодированы и некоторые белки, входящие в состав рибосом этих органелл. Ключевые ферменты матричного полинуклеоти-дного синтеза, такие как ДНК-полимераза (осуществляющая репликацию мито-хондриальной ДНК) и РНК-полимераза (транскрибирующая геном митохон-дрий), зашифрованы в ядре и синтезируются на рибосомах цитоплазмы. Этот факт указывает на относительность автономии митохондрий в сложной иерар-хии эвкариотической клетки.
Геномы митохондрий разных видов отличаются не только по набору ге-нов, порядку их расположения и экспрессии, но по размеру и форме ДНК. По-давляющее большинство описанных сегодня митохондриальных геномов пред-ставляет собой кольцевые суперспирализованные двуцепочечные молекулы ДНК. У некоторых растений наряду с кольцевыми формами имеются и линей-ные, а у некоторых простейших, например инфузорий, в митохондриях обнару-жены только линейные ДНК.
Как правило, в каждой митохондрии содержится несколько копий ее ге-нома. Так, в клетках печени человека около 2 тыс. митохондрий, и в каждой из них — по 10 одинаковых геномов. В фибробластах мыши 500 митохондрий, со-держащих по два генома, а в клетках дрожжей S.cerevisiae — до 22 митохон-дрий, имеющих по четыре генома.
Митохондриальный геном растений, как правило, состоит из нескольких молекул разного размера. Одна из них, “основная хромосома”, содержит боль-шую часть генов, а кольцевые формы меньшей длины, находящиеся в динами-ческом равновесии как между собой, так и с основной хромосомой, образуются в результате внутри- и межмолекулярной рекомбинации благодаря наличию по-вторенных последовательностей (рис.1).
Рис 1. Схема образования кольцевых молекул ДНК разного размера в митохондриях растений. Рекомбинация происходит по повторенным участкам (обозначены синим цветом).
В митохондриях большинства организмов (кроме высших животных) часть кольцевых молекул ДНК присутствует в виде олигоме-ров, которые можно разделить на три класса: линейные; кольцевые, имеющие контурную длину, кратную длине мономерных колец; цепные, катенаны, состо-ящие из топологически связанных, т.е. продетых друг в друга, мономерных ко-лец (рис.2). Так, в единственной митохондрии простейших из отряда кинето-пластид, включающего эндопаразита человека — трипаносому, содержатся ты-сячи кольцевых молекул ДНК. У Trypanosoma brucei имеются два типа моле-кул: 45 одинаковых максиколец, каждое из которых состоит из 21 тыс. пар ну-клеотидов, и 5.5 тыс. идентичных друг другу миниколец по 1000 пар нуклео-тидов. Все они, соединяясь в катенаны, образуют переплетенную сеть, которая вместе с белками формирует структуру, называемую кинетопластом.
 Рис
2. Схема образования линейных (А), кольцевых (Б), цепных (В) олигомеров
мтДНК. ori — район начала репликации ДНК.
Рис
2. Схема образования линейных (А), кольцевых (Б), цепных (В) олигомеров
мтДНК. ori — район начала репликации ДНК.
Размер генома митохондрий разных организмов колеблется от менее 6 тыс. пар нуклеотидов у малярийного плазмодия (в нем, помимо двух генов рРНК, содержится только три гена, кодирующих белки) до сотен тысяч пар ну-клеотидов у наземных растений (например, у Arabidopsis thaliana из семейства крестоцветных 366924 пар нуклеотидов). При этом 7—8-кратные различия в ра-змерах мтДНК высших растений обнаруживаются даже в пределах одного се-мейства. Длина мтДНК позвоночных животных отличается незначительно: у человека — 16569 пар нуклеотидов, у свиньи — 16350, у дельфина — 16330, у шпорцевой лягушки Xenopus laevis — 17533, у карпа — 16400. Эти геномы схо-дны также и по локализации генов, большинство которых располагаются встык; в ряде случаев они даже перекрываются, обычно на один нуклеотид, так что по-следний нуклеотид одного гена оказывается первым в следующем. В отличие от позвоночных, у растений, грибов и простейших мтДНК содержат до 80% не-кодирующих последовательностей. У разных видов порядок генов в геномах митохондрий отличается.
Высокая концентрация активных форм кислорода в митохондриях и сла-бая система репарации увеличивают частоту мутаций мтДНК по сравнению с ядерной на порядок. Радикалы кислорода служат причиной специфических за-мен Ц®Т (дезаминирование цитозина) и Г®Т (окислительное повреждение гуанина), вследствие чего, возможно, мтДНК богаты АТ-парами. Кроме того, все мтДНК обладают интересным свойством — они не метилируются, в отли-чие от ядерных и прокариотических ДНК. Известно, что метилирование (време-нная химическая модификация нуклеотидной последовательности без наруше-ния кодирующей функции ДНК) — один из механизмов программируемой инактивации генов.
Размеры и строение молекул ДНК в органеллах
|
Вид |
Структура |
Масса, млн. дальтон |
Примечания |
|
|
Мит охон дриа льн ая Д Н К |
Животные |
Кольцевая | 9-12 | У каждого отдельного вида все молекулы одного размера |
|
Высшие ра стения |
Кольцевая | Варьирует | У всех изученных видов имеются разные по величине кольцевые ДНК, в которых общее содержание генетической информации соответ-ствует массе от 300 до 1000 млн. дальтон в зависимости от вида | |
|
Грибы: Saccharomyces Kluyveromyces Простейшие Plasmodium Paramecium |
Кольцевая Кольцевая Кольцевая Линейная |
50 22 18 27 |
||
|
Д Н К Хлор опла стов |
Водоросли Chlamydomonas Euglena |
Кольцевая Кольцевая |
120 90 |
|
|
Высшие растения |
Кольцевая | 85-97 |
У каждого отдельного вида найдены молекулы только одного размера |
|
Относительное количество ДНК органелл в некоторых клетках и тканях
|
Организм |
Ткань или тип клеток |
Число мол-л ДНК/органел- лу |
Число орга- нелл в клетке |
Доля ДНК орга-нелл во всей ДНК клетки, % |
|
|
Мит охон дриа льн ая Д Н К |
Крыса | Печень | 5-10 | 1000 | 1 |
| Мышь | Клетки линии L | 5-10 | 100 | <1 | |
| Лягушка | Яйцеклетка | 5-10 |
107 |
99 | |
| Дрожжи | Вегетативные диплоидные клетки | 2-50 | 2-50 | 15 | |
|
Д Н К Хлор опла стов |
Chlamydomonas | Вегетативные диплоидные клетки | 80 |
2
|
7 |
| Кукуруза | Листья | 20-40 | 20-40 | 15 | |
Функционирование митохондриального генома
Что же особенного в механизмах репликации и транскрипции ДНК митохондрий млекопитающих?
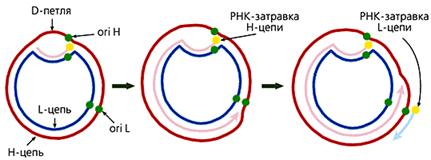 У большинства животных
комплементарные цепи в мтДНК значительно различаются по удельной плотности,
поскольку содержат неодинаковое количе-ство “тяжелых” пуриновых и “легких”
пиримидиновых нуклеотидов. Так они и называются — H (heavy — тяжелая) и L
(light — легкая) цепь. В начале репли-кации молекулы мтДНК образуется так
называемая D-петля (от англ. Displace-ment loop — петля смещения). Эта
структура, видимая в электронный микро-скоп, состоит из двуцепочечного и
одноцепочечного (отодвинутой части Н-цепи) участков. Двуцепочечный участок
формируется частью L-цепи и компле-ментарным ей вновь синтезированным
фрагментом ДНК длиной 450—650 (в зависимости от вида организма) нуклеотидов,
имеющим на 5'-конце рибонук-леотидную затравку, которая соответствует точке
начала синтеза Н-цепи (oriH). Синтез L-цепи начинается лишь тогда, когда
дочерняя Н-цепь доходит до точки ori L. Это обусловлено тем, что область
инициации репликации L-цепи доступ-на для ферментов синтеза ДНК лишь в
одноцепочечном состоянии, а следовате-льно, только в расплетенной двойной
спирали при синтезе Н-цепи. Таким обра-зом, дочерние цепи мтДНК синтезируются
непрерывно и асинхронно (рис.3).
У большинства животных
комплементарные цепи в мтДНК значительно различаются по удельной плотности,
поскольку содержат неодинаковое количе-ство “тяжелых” пуриновых и “легких”
пиримидиновых нуклеотидов. Так они и называются — H (heavy — тяжелая) и L
(light — легкая) цепь. В начале репли-кации молекулы мтДНК образуется так
называемая D-петля (от англ. Displace-ment loop — петля смещения). Эта
структура, видимая в электронный микро-скоп, состоит из двуцепочечного и
одноцепочечного (отодвинутой части Н-цепи) участков. Двуцепочечный участок
формируется частью L-цепи и компле-ментарным ей вновь синтезированным
фрагментом ДНК длиной 450—650 (в зависимости от вида организма) нуклеотидов,
имеющим на 5'-конце рибонук-леотидную затравку, которая соответствует точке
начала синтеза Н-цепи (oriH). Синтез L-цепи начинается лишь тогда, когда
дочерняя Н-цепь доходит до точки ori L. Это обусловлено тем, что область
инициации репликации L-цепи доступ-на для ферментов синтеза ДНК лишь в
одноцепочечном состоянии, а следовате-льно, только в расплетенной двойной
спирали при синтезе Н-цепи. Таким обра-зом, дочерние цепи мтДНК синтезируются
непрерывно и асинхронно (рис.3).
Рис 3. Схема репликации мтДНК млекопитающих. Сначала формируется D-петля, затем синтезируется дочерняя Н-цепь, потом начинается синтез дочерней L-цепи.
 В митохондриях общее число молекул с
D-петлей значительно превыша-ет число полностью реплицирующихся молекул.
Обусловлено это тем, что у D-петли есть дополнительные функции — прикрепление
мтДНК к внутренней ме-мбране и инициация транскрипции, поскольку в этом районе
локализованы промоторы транскрипции обеих цепей ДНК. В отличие от большинства
эв-кариотических генов, которые транскрибируются независимо друг от друга, ка-ждая
из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК,
начинающейся в районе ori H. Помимо этих двух длинных мо-лекул РНК,
комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые
начинаются в той же точке и заканчиваются на 3'-кон-це гена 16S рРНК (рис.4).
Таких коротких транскриптов в 10 раз больше, чем длинных. В результате
созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в
формировании митохондриальных рибосом, а так-же фенилаланиновая и валиновая
тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются
транслируемые мРНК, к 3'-концам которых при-соединяются полиадениловые
последовательности. 5'-концы этих мРНК не кэ-пируются, что необычно для
эвкариот. Сплайсинга (сращивания) не происхо-дит, поскольку ни один из
митохондриальных генов млекопитающих не содер-жит интронов.
В митохондриях общее число молекул с
D-петлей значительно превыша-ет число полностью реплицирующихся молекул.
Обусловлено это тем, что у D-петли есть дополнительные функции — прикрепление
мтДНК к внутренней ме-мбране и инициация транскрипции, поскольку в этом районе
локализованы промоторы транскрипции обеих цепей ДНК. В отличие от большинства
эв-кариотических генов, которые транскрибируются независимо друг от друга, ка-ждая
из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК,
начинающейся в районе ori H. Помимо этих двух длинных мо-лекул РНК,
комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые
начинаются в той же точке и заканчиваются на 3'-кон-це гена 16S рРНК (рис.4).
Таких коротких транскриптов в 10 раз больше, чем длинных. В результате
созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в
формировании митохондриальных рибосом, а так-же фенилаланиновая и валиновая
тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются
транслируемые мРНК, к 3'-концам которых при-соединяются полиадениловые
последовательности. 5'-концы этих мРНК не кэ-пируются, что необычно для
эвкариот. Сплайсинга (сращивания) не происхо-дит, поскольку ни один из
митохондриальных генов млекопитающих не содер-жит интронов.
|
ND1—ND6, ND4L — гены субъединиц НAД-H-дегидрогеназного комплекса; СОI—COIII — гены субъединиц цитохром-с-оксидазы; ATP6, ATP8 — гены субъединиц AТФ-синтетазы Cyt b — ген цитохрома b. |
Рис 4. Транскрипция мтДНК человека, содержащей 37 генов. Все транскрипты начинают синтезироваться в районе ori H. Рибосомные РНК вырезаются из длинного и короткого транскриптов Н-цепи. тРНК и мРНК образуются в результате процессинга из транскриптов обеих цепей ДНК. Гены тРНК обозначены светло-зеленым цветом.
Хотите узнать какие еще сюрпризы способен преподнести митохон-дриальный геном? Отлично! Читаем дальше!..
Несмотря на то, что в геномах митохондрий млекопитающих и дрожжей содержится приблизительно одинаковое количество генов, размеры дрожжево-го генома в 4-5 раз больше — около 80 тыс. пар нуклеотидов. Хотя кодирую-щие последовательности мтДНК дрожжей высоко гомологичны соответствую-щим последовательностям у человека, дрожжевые мРНК дополнительно имеют 5'-лидерную и 3'-некодирующую области, как и большинство ядерных мРНК. Ряд генов содержит еще и интроны. Так, в гене box, кодирующем цитохром-оксидазу b, имеется два интрона. Из первичного РНК-транскрипта автокатали-тически (без участия каких-либо белков) вырезается копия большей части пер-вого интрона. Оставшаяся РНК служит матрицей для образования фермента ма-туразы, участвующей в сплайсинге. Часть ее аминокислотной последовательно-сти закодирована в оставшихся копиях интронов. Матураза вырезает их, разру-шая свою собственную мРНК, копии экзонов сшиваются, и образуется мРНК для цитохромоксидазы b (рис.5). Открытие такого феномена заставило пере-смотреть представление об интронах, как о “ничего не кодирующих последова-тельностях”.
Рис 5. Процессинг (созревание) мРНК цитохромоксидазы b в митохондриях дрожжей. На первом этапе сплайсинга образуется мРНК, по которой синтезируется матураза, необходимая для второго этапа сплайсинга.
При изучении экспрессии митохон-дриальных генов Trypanosoma brucei
обнаружилось удивительное отклонение от одной из основных аксиом молекулярной
биологии, гласящей, что после-довательность нуклеотидов в мРНК в точности
соответствует таковой в коди-рующих участках ДНК. Оказалось, мРНК одной из
субъединиц цитохром-с-оксидазы редактируется,
т.е. после транскрипции изменяется ее первичная структура — вставляется четыре
урацила. В результате образуется новая мРНК, служащая матрицей для синтеза
дополнительной субъединицы фермента, пос-ледовательность аминокислот в которой
не имеет ничего общего с последова- тельностью, кодируемой
нередактированной мРНК (см. таблицу).
тельностью, кодируемой
нередактированной мРНК (см. таблицу).
