

 Доклад: Неканонические наследственные изменения
Доклад: Неканонические наследственные изменения
Открытия в области подвижной генетики показали, что клетка как целостная система в ходе отбора может адаптивно перестраивать свой геном. Она способна ответить на вызов среды активным генетическим поиском, а не пассивно ждать случайного возникновения мутации, позволяющей выжить. А в опытах супругов Ледерберг у клеток не было выбора: либо смерть, либо адаптивная мутация.
В тех же случаях, когда фактор отбора не летален, возможны постепенные перестройки генома, прямо или косвенно связанные с условиями отбора. Это выяснилось с открытием в конце 70-х годов постепенного умножения числа локусов, в которых расположены гены устойчивости к селективному агенту, блокирующему деление клеток. Известно, что метотрексат — ингибитор клеточного деления — широко применяется в медицине для остановки роста злокачественных клеток. Этот клеточный яд инактивирует фермент дигидрофолатредуктазу (ДГФР), работу которого контролирует определенный ген.
Рассмотрим, как в ходе подобного отбора менялся геном у одноклеточного паразитического жгутиконосца лейшмании Leishmania tropica, вызывающего кожные язвы и передающегося человеку москитами от грызунов. До начала опытов ген устойчивости был в геноме лейшмании в одной дозе. В результате отбора число генов устойчивости увеличилось (амплифицировалось). Что же наблюдали при разных вариантах отбора?
Устойчивость клеток лейшмании к яду-цитостатику (метотрексат) возрастала ступенчато, и пропорционально увеличивалась доля амплифицированных сегментов с геном устойчивости. Умножался не только селектируемый ген, но и большие прилежащие к нему участки ДНК, названные ампликонами. Когда устойчивость к яду у лейшмании повысилась в 1000 раз, амплифицированные внехромосомные сегменты составили до 10% ДНК в клетке! Можно сказать, что из одного облигатного гена образовался пул факультативных элементов. Произошла адаптивная перестройка генома в ходе отбора.
Если отбор продолжался достаточно долго, часть ампликонов встраивалась в исходную хромосому, и после прекращения отбора устойчиво сохранялась повышенная устойчивость.
С удалением из среды селективного агента число ампликонов с геном устойчивости постепенно снижалось в ряду поколений и одновременно падала устойчивость. Тем самым был смоделирован феномен длительных модификаций, когда массовые изменения, вызванные средой, наследуются, но постепенно угасают в ряду поколений.
При повторном отборе часть сохранившихся в цитоплазме ампликонов обеспечивала быструю их автономную репликацию, и устойчивость возникала гораздо быстрее, чем в начале опытов. Иными словами, формировалась своеобразная клеточная ампликонная память о прошедшем отборе на основе сохранившихся ампликонов.
Если сопоставить метод реплик и ход отбора на устойчивость в случае амплификации, то оказывается, что именно контакт с селективным фактором вызвал преобразование генома, характер которого коррелировал с интенсивностью и направлением отбора.
Дискуссия об адаптивных мутациях
В 1988 г. в журнале “Nature” появилась статья Дж.Кэйрнса с соавторами о возникновении у бактерии E.coli отборзависимых “направленных мутаций”. Брали бактерии, несущие мутации в гене lacZ лактозного оперона, неспособные расщеплять дисахарид лактозу. Но эти мутанты могли делиться на среде с глюкозой, откуда их через один—два дня роста переносили на селективную среду с лактозой. Отобрав lac+ реверсов, которые, как и ожидалось, возникли еще в ходе “глюкозных” делений, нерастущие клетки оставляли в условиях углеводного голодания. Сначала мутанты отмирали. Но спустя неделю и более наблюдался новый рост за счет вспышки реверсий именно в гене lacZ. Как будто клетки в условиях жесткого стресса, не делясь (!), вели генетический поиск и адаптивно меняли свой геном [10].
В последующих работах Б.Холла использовались бактерии, мутантные по гену утилизации триптофана (trp). Их помещали на среду, лишенную триптофана, и оценивали частоту реверсий к норме, которая повышалась именно при триптофановом голодании. Но причиной этого феномена были не сами условия голодания, ибо на среде с голоданием по цистеину частота реверсий к trp+ не отличалась от нормы.
В следующей серии опытов Холл взял уже двойных недостаточных по триптофану мутантов, несущих одновременно мутации в генах trpA и trpВ, и вновь поместил бактерии на среду, лишенную триптофана. Выжить могли только особи, у которых реверсии возникали одновременно в двух триптофановых генах. Частота появления таких особей была в 100 млн раз выше, чем ожидалось при простом вероятностном совпадении мутаций в двух генах. Холл предпочел называть этот феномен “адаптивные мутации” и впоследствии показал, что они возникают и у дрожжей, т.е. у эвкариот [11].
Публикации Кэйрнса и Холла немедленно вызвали бурную дискуссию. Итогом ее первого раунда стало выступление одного из ведущих исследователей в области подвижной генетики Дж.Шапиро. Он кратко обсудил две основные идеи. Во-первых, клетка содержит биохимические комплексы, или системы “естественной генетической инженерии”, которые способны реконструировать геном. Активность этих комплексов, как и любая клеточная функция, может резко меняться в зависимости от физиологии клетки. Во-вторых, частота возникновения наследственных изменений всегда оценивается не для одной клетки, а для клеточной популяции, в которой клетки могут обмениваться между собой наследственной информацией. Кроме того, межклеточный горизонтальный перенос с помощью вирусов или передачи сегментов ДНК усиливается в стрессовых условиях. Как считает Шапиро, эти два механизма объясняют феномен адаптивных мутаций и возвращают его в русло обычной молекулярной генетики [11]. Каковы же, на его взгляд, итоги дискуссии? “Мы нашли там генетического инженера с впечатляющим набором замысловатых молекулярных инструментов для реорганизации ДНК-молекулы” [12].
За последние десятилетия на уровне клетки открыта такая непредвиденная сфера сложности и координации, которая более совместима с компьютерной технологией, нежели с механизированным подходом, доминировавшим во время создания неодарвинистского современного синтеза. Вслед за Шапиро, можно назвать по крайней мере четыре группы открытий, изменивших понимание клеточных биологических процессов.
Организация генома. У эвкариот генетические локусы устроены по модульному принципу, представляя собой конструкции из регуляторных и кодирующих модулей, общих для всего генома. Это обеспечивает быструю сборку новых конструкций и регуляцию генных ансамблей. Локусы организованы в иерархические сети, во главе с главным геном-переключателем (как в случае регуляции пола или развития глаза). Причем многие из соподчиненных генов интегрированы в разные сети: они функционируют в разные периоды развития и влияют на множество признаков фенотипа.
Репаративные возможности клетки. Клетки вовсе не пассивные жертвы случайных физико-химических воздействий, поскольку в них имеется система репараций на уровне репликации, транскрипции и трансляции.
Мобильные генетические элементы и природная генетическая инженерия. Работа иммунной системы построена на непрерывном конструировании новых вариантов молекул иммуноглобулинов на основе действия природных биотехнологических систем (ферменты: нуклеазы, лигазы, обратные транскриптазы, полимеразы и т.д.). Эти же системы используют мобильные элементы для создания новых наследуемых структур. При этом генетические изменения могут быть массовыми и упорядоченными. Реорганизация генома — один из основных биологических процессов. Природные генноинженерные системы регулируются системами с обратной связью. До поры до времени они пребывают в неактивном состоянии, но в ключевые периоды или во время стресса приводятся в действие.
Клеточный информационный процессинг. Возможно, одно из самых важных открытий в области биологии клетки состоит в том, что клетка непрерывно собирает и анализирует информацию о своем внутреннем состоянии и внешней среде, принимая решение о росте, движении и дифференциации. Особенно показательны механизмы контроля клеточного деления, лежащие в основе роста и развития. Процесс митоза универсален у высших организмов и включает три последовательных этапа: подготовка к делению, репликация хромосом и завершение деления клетки. Анализ генного контроля этих фаз привел к открытию особых точек, в которых клетка проверяет, произошла ли репарация нарушений в структуре ДНК на предыдущем этапе или нет. Если ошибки не будут исправлены, последующий этап не начнется. Когда же ликвидировать повреждения нельзя, запускается генетически запрограммированная система клеточной смерти, или апоптоза.
В условиях вызова среды клетка действует целенаправленно, подобно компьютеру, когда при его запуске шаг за шагом проверяется нормальная работа основных программ, и в случае неисправности работа компьютера останавливается. В целом становится очевидной, уже на уровне клетки, правота нетрадиционного французского зоолога-эволюциониста Поля Грассэ: “Жить — значит реагировать, а отнюдь не быть жертвой”.
Пути возникновения естественных наследственных изменений в системе среда—факультативные элементы—облигатные элементы. Факультативные элементы первыми воспринимают немутагенные факторы среды, а возникающие затем вариации вызывают мутации. На поведение факультативных элементов влияют и облигатные элементы.
Неканонические наследственные изменения, возникающие под влиянием отбора к цитостатикам и приводящие к амплификации генов.
Приобретенные признаки наследуются
“История биологии не знает более выразительного примера многовекового обсуждения проблемы, чем дискуссия о наследовании или о ненаследовании приобретенных признаков”, - эти слова стоят в начале книги известного цитолога и историка биологии Л.Я.Бляхера [12]. В истории, пожалуй, можно вспомнить аналогичную ситуацию с попытками превращения химических элементов. Алхимики верили в эту возможность, но в химии утвердился постулат о неизменности химических элементов. Однако ныне в ядерной физике и химии исследования по превращению элементов и анализ их эволюции - дело обычное. Кто же оказался прав в многовековом споре? Можно сказать, что на уровне химических молекулярных взаимодействий не происходит превращения элементов, а на ядерном уровне оно - правило.
Напрашивается сходная аналогия и с вопросом о наследовании признаков, которые появились в ходе онтогенеза. Если вновь возникающие наследственные изменения сводить лишь к мутациям генов и хромосом, тогда вопрос можно считать закрытым. Но если исходить из обобщенной концепции генома, включая представление о динамической наследственности [2, 3], проблема нуждается в пересмотре. Помимо мутационной существует вариационная и эпигенетическая формы наследственной изменчивости, связанные не с изменениями в тексте ДНК, а в состоянии гена. Такие эффекты обратимы и наследуемы.
Интересно, что вышедший в конце 1991 г. Международный ежегодник по генетике открывается статьей О.Ландмана “Наследование приобретенных признаков” [13]. Автор суммирует уже давно полученные в генетике факты, показывая, что “наследование приобретенных признаков вполне совместимо с современной концепцией молекулярной генетики”. Ландман детально рассматривает около десяти экспериментальных систем, в которых установлено наследование приобретенных признаков. Четыре разных механизма способны привести к нему: изменение структур клеточной оболочки, или кортекса, изученное Т. Соннеборном у инфузорий; ДНК-модификации, т.е. клонально передаваемые изменения в характере локального метилирования ДНК (сюда входит феномен импринтинга); эпигенетические изменения без каких-либо модификаций ДНК; индуцированная утрата либо приобретение факультативных элементов.
Статья Ландмана делает нас как бы свидетелями критического периода смены постулата в генетике, казавшегося неколебимым как скала. Автор спокойно, без ажиотажа и новых сногсшибательных фактов, объединяет старые и новые данные в систему, дает им ясное современное истолкование. Можно сформулировать общий принцип: наследование приобретенных признаков возможно в тех случаях, когда некий фенотипический признак зависит от числа или топографии факультативных элементов.
Приведу два поучительных примера на дрозофиле: первый связан с поведением вируса сигма, второй - мобильных элементов, ответственных за гибридную стерильность самок и сверхмутабильность.
Изучение взаимодействия вируса сигма с геномом дрозофилы началось более 60 лет назад. Сначала в 1937 г. французский генетик Ф.Леритье обнаружил резкие наследственные отличия у разных линий мух по степени чувствительности к углекислому газу (СО2). Признак наследовался причудливым образом: через цитоплазму, но не только по материнской линии, а иногда и через самцов. Чувствительность можно было передать и путем инъекции гемолимфы, причем разным видам дрозофил. В этих случаях признак передавался не стабильно, но в результате отбора наследование становилось устойчивым.
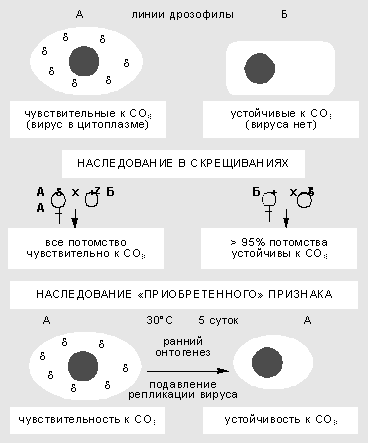
Неменделевское наследование признака у дрозофилы, который зависит от популяции факультативных элементов генома. Признак чувствительности к СО2 вызван присутствием в цитоплазме мухи рабдовируса сигма. В результате температурного шока на ранней стадии развития дрозофилы блокируется размножение вируса, и выросшие особи приобретают устойчивость к нему.
Чувствительность к СО2 оказалась связана с устойчивым размножением в половых и соматических клетках РНК-содержащего пулевидного рабдовируса сигма, сходного по ряду свойств с вирусом бешенства у млекопитающих. Оогонии (клетки, из которых в ходе мейоза и созревания образуются яйцеклетки) у самок стабилизированной линии обычно содержат 10-40 вирусных частиц, а ооциты (зрелые яйцеклетки) - 1-10 млн. Вирус сигма - типичный факультативный элемент. Мутации в его геноме приводят к сложным формам поведения системы. Найдены случаи вирусоносительства, при которых дрозофилы остаются устойчивы к СО2, но вместе с тем иммунны к заражению другими штаммами вируса. Ситуация вполне сравнима с поведением системы фаг-бактерия, что сразу заметили Ф.Жакоб и Э.Вольман.
Взаимоотношения генома дрозофилы и размножающегося в ее цитоплазме вируса подчиняются правилам внутриклеточной генетики. Воздействия в ходе онтогенеза могут вызвать сдвиг в числе и межклеточной топографии частиц и как результат - изменить степень чувствительности к углекислому газу. Так, повышенная температура блокирует репликацию вирусных частиц. Если самок и самцов в период гаметогенеза содержать несколько дней при температуре 30°С, потомство от таких мух будет свободно от вируса и устойчиво к СО2. Значит, приобретенный в ходе индивидуального развития признак наследуется в ряду поколений.
Ситуация с вирусом сигма не единична. Французские генетики изучали факторы стерильности самок, связанные с поведением мобильных элементов типа “I”. Наследование этого признака определяется сложными ядерно-цитоплазматическими взаимодействиями. Если в отцовских хромосомах локализованы активные I-элементы, то на фоне R-цитоплазмы они начинают активироваться, претерпевают множественные транспозиции и в результате вызывают резкие нарушения онтогенеза в потомстве самок с чувствительной цитоплазмой. Такие самки откладывают яйца, но часть эмбрионов гибнет на ранней стадии дробления - еще до образования бластомеры. Линии, выделенные из природных популяций, отличаются по силе действия I-факторов и степени реактивности (или чувствительности) цитоплазмы. Эти показатели можно изменить внешним влиянием. Возраст исходных родительских самок, а также воздействие в ранний период развития повышенной температуры отражаются не только на плодовитости выросших самок, но и на плодовитости их потомства. Вызванные условиями среды изменения реактивности цитоплазмы поддерживаются на протяжении многих клеточных поколений. “Самое замечательное, что эти изменения реактивности цитоплазмы под влиянием негенетических факторов наследуются: наблюдается наследование “благоприобретенных” признаков”, - отмечал Р.Б.Хесин [5].
Наследование через цитоплазму: от бабушек до внуков
В теории развития и феногенетике ХХ в. важное место занимают глубокие и совершенно оригинальные исследования эмбриолога П.Г.Светлова (1892-1972). Остановимся на разработанной им теории квантованности онтогенеза (наличии критических периодов в развитии, когда происходит детерминация морфогенетических процессов и одновременно повышается чувствительность клеток к повреждающим агентам) и на развитой в связи с этим идее, что изучение онтогенеза надо вести не с момента оплодотворения и образования зиготы, а еще с гаметогенеза, включающего оогенез у самок предшествующего поколения - проэмбрионального периода.
